A rethorical question, obviously, with an equally obvious, trivial answer: Well, it depends…
Depends on what, however, is less trivial, and deserves a closer look at the evidence available.
Let me start with distinguishing Colonisation from male-driven genetic replacement. The former entails a joint migration of males and females into new territories, e.g. the European settlement of the USA and Canada, or, further back in time, colonisation of much of Europe by Near Eastern Farmers. Colonisation typically leads to language-shift, if the colonisers have sufficient demographic and/or socio-cultural weight to overpover the “natives”, as well demonstrated for the Americas and Australia. In that sense, there is every reason to assume that EEF brought with them new languages to the parts of Europe they colonised, linked to either the Levante or Anatolia [I plan for a separate posting on EEF languages]. Without sufficient such weight, OTOH, colonisers may have failed to excert substantial linguistic impact. Points in case are the Vandals in NW Africa, and more generally various early medieval Germanics migrating into the Roman Empire (Visigoths, Lombards etc.), also antique Greeks west of S. Italy / Sicily, or in Bactria.
In contrast, male-driven genetic replacement is assymetrical, i.e. relating mostly to yDNA but hardly impacting mtDNA. In those good-old, pre autosomal DNA times not so long ago, we (or at least I myself) looked for possible associations between yDNA and languages. The problems of that approach have become fairly obvious by now: yDNA N1c, once firmly believed to be an Uralic marker, has in the meantime been demonstrated to have been associated with the Northern Chinese MN (after ca. 4500 BC) that is unlikely to have spoken anything akin to Proto-Uralic, while, aside from a stray find in 3rd mBC Comb Ceramic contexts near Smolensk, connection of N1c to Proto-Uralic still needs to be demonstrated. R1b, traditionally either seen as a Western IE (Germanic-Celtic) or Basque marquer, and, in its V88 variant typical of Afro-Asiatic (Chadic/ Berber)-speaking populations, is by now earliest attested from an epi-Paleolithic WHG context (Vilabruna) that is neither likely to have spoken anything akin to PIE, nor to early Afro-Asiatic.
Nevertheless, it isn’t unreasonable at all to assume that male-driven genetic replacement may induce language shift. Strong evidence comes from Africa, where an older study (forgot the source, but surely one of the readers/ commentators can help out in this respect) demonstrated an assymetrical relation of male and female DNA. While variance of the latter was closely connected to geography (isolation-by-distance), the former rather followed linguistic separation. Such a macro-analysis is obviously not able to fully capture the genetic and linguistic history of the whole continent. More specifically, issues like the following certainly deserve further exploration:
- Reasons for the persistence of certain, apparently very ancient HG languages like Hazda and members of the Khoi-San family (the latter with demonstrated Semitic genetic influence);
- A closer look at the genesis of certain Afro-Asiatic languages, especially Berber, against the background of Iberomaurusian aDNA;
- Zooming in on Madagascar with its specific, genetic as well as linguistic, mix of ancient HGs, Bantu, Austronesians, and Near Easterners (yDNA T, 60% among aristocratic Anteony);
- the history of Nilo-Saharan, which, while culturally hardly distinguishable from Bantu- and Chadic-speakers, nevertheless seems to have been partly overpowered by both.
Still, at least as concerns speakers of Chadic and Bantu languages, genetic analysis aligns with other (linguistic, archeological) data pointing towards a massive expansion from ca. 1,000 BC onwards that was male-driven, rooted in a (semi-) nomadic, pastoralist lifestyle, and, while incorporating females from the territories overtaken, mostly replaced HG yDNA there. Importantly, the pastoralists also introduced new linguae francae such as Suahili (Bantu) or Hausa (Chadic). Of course, since times immemorible, nomadic pastoralists have also been traders, as illustrated by Trans-Saharan caravans, going back to camel herders exploring a specific ecologic niche, and initially carrying back just some salt to sell to cattle herders in the Sahel, before finding out that there was much more to trade with if they continued their trail towards Morocco, Lybia or Egypt.
So far, so good. Now comes, for another part of the world, C. Posth e.a. “Language continuity despite population replacement in Remote Oceania” (full publication, paywalled, here):
 “Recent genomic analyses show that the earliest peoples reaching Remote Oceania—associated with Austronesian-speaking Lapita culture—were almost completely East Asian, without detectable Papuan ancestry. However, Papuan-related genetic ancestry is found across present-day Pacific populations, indicating that peoples from Near Oceania have played a significant, but largely unknown, ancestral role. Here, new genome-wide data from 19 ancient South Pacific individuals provide direct evidence of a so-far undescribed Papuan expansion into Remote Oceania starting ~2,500 yr bp, far earlier than previously estimated and supporting a model from historical linguistics. New genome-wide data from 27 contemporary ni-Vanuatu demonstrate a subsequent and almost complete replacement of Lapita-Austronesian by Near Oceanian ancestry. Despite this massive demographic change, incoming Papuan languages did not replace Austronesian languages. Population replacement with language continuity is extremely rare—if not unprecedented—in human history. Our analyses show that rather than one large-scale event, the process was incremental and complex, with repeated migrations and sex-biased admixture with peoples from the Bismarck Archipelago.”
“Recent genomic analyses show that the earliest peoples reaching Remote Oceania—associated with Austronesian-speaking Lapita culture—were almost completely East Asian, without detectable Papuan ancestry. However, Papuan-related genetic ancestry is found across present-day Pacific populations, indicating that peoples from Near Oceania have played a significant, but largely unknown, ancestral role. Here, new genome-wide data from 19 ancient South Pacific individuals provide direct evidence of a so-far undescribed Papuan expansion into Remote Oceania starting ~2,500 yr bp, far earlier than previously estimated and supporting a model from historical linguistics. New genome-wide data from 27 contemporary ni-Vanuatu demonstrate a subsequent and almost complete replacement of Lapita-Austronesian by Near Oceanian ancestry. Despite this massive demographic change, incoming Papuan languages did not replace Austronesian languages. Population replacement with language continuity is extremely rare—if not unprecedented—in human history. Our analyses show that rather than one large-scale event, the process was incremental and complex, with repeated migrations and sex-biased admixture with peoples from the Bismarck Archipelago.”
The sex-biased admixture described relates to Papuan males intermixing with Austronesian women, now for the first time directly documented from aDNA, but already long assumed based on uniparental markers. Especially Polynesian, to a lesser extent also Melanesian and Insular SEA mtDNA is dominated by hg B4a1a1, the so-called “Polynesian motif”. Upstream B4a1a is quite frequent on the Phillipines, and accounts for 23% of the mtDNA of Austronesian-speaking Ami from Taiwan (source, Tab. S8) Its connection to the spread of Austronesian is a/o documented through the B4a1a1a2 subclade that is dominating among Malagassy. In contrast, Polynesian and Melanesian yDNA is far more diverse, including (in distributions that may vary widely from archipelago to archipelago) hgs C-M38 (formerly C2), K, M and S. This phenomenon – mitochochondrial and linguistic uniformity, but paternal diversity – has traditionally been explained from the regions’ matriarchic/ matrilocal culture. The Posth paper points add another possible reason, namely Austronesian acting as lingua franca over a wide region (today, Austronesian is spoken from Madagascar in the West to Hawaii in the East and New Zealand in the South).
Before further exploring the main question, allow me to briefly stay in the region to point at another unusal case of language shift, namely Australian Pama-Nyungan languages. Covering 90% of the continent, and comprising up to 400 languages prior to European settlement that commenced 250 years ago, this used to be one of the world’s largest language families in terms of diversity and geographical spread.
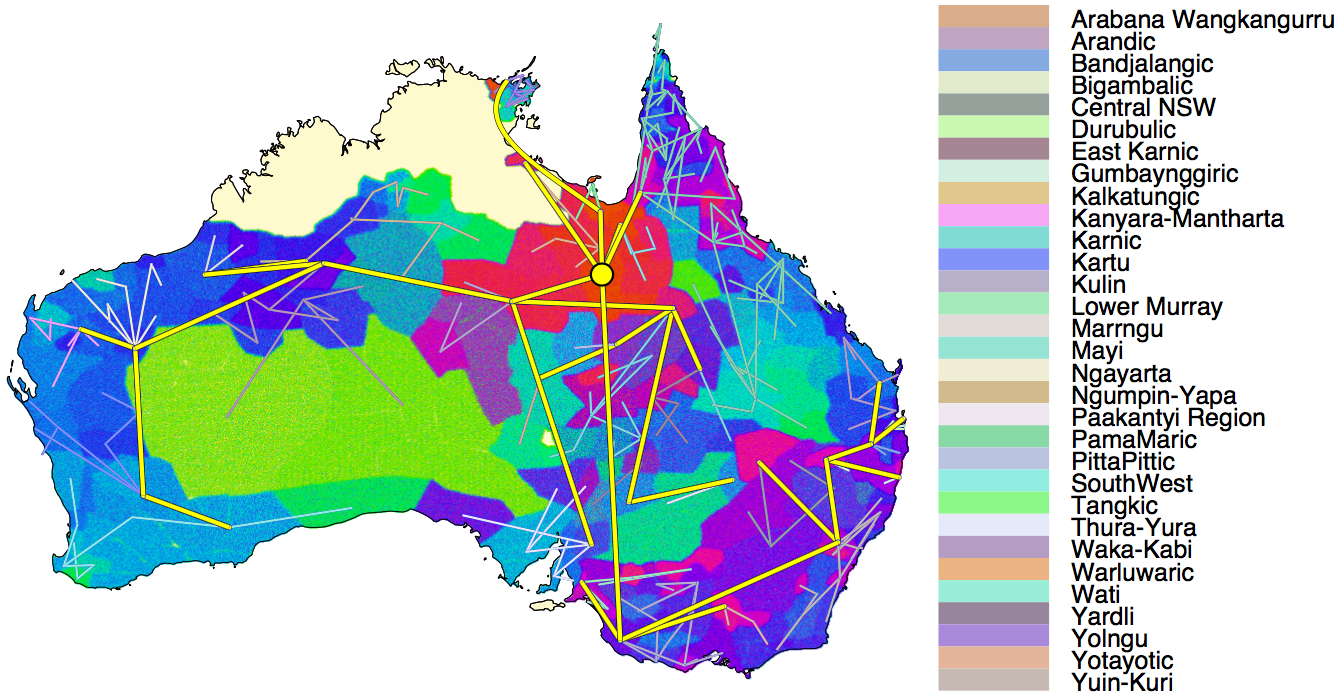
Recently, J. Bouckaert e.a. combined new linguistic data on 306 Pama-Nyungan languages with a set of modelling tools initially developed by epidemiologists to trace the origin of virus outbreaks. They constructed a phylogenic language tree based on Bayesian modelling, from which they inferred the root age and homeland of Proto-Pama-Nyungan, and traced the family’s expansion through time and space. Their approach and findings have been published in Nature (pay-walled), and are presented and discussed quite extensively on a dedicated home-page (link). Their analyses “support an origin of Pama-Nyungan 5,000 to 7,000 years ago originating from just south of the Gulf of Carpentaria, consistent with [a scenario of] rapid replacement [of preceding languages]“; a scenario “linked in the literature to one or more of technological advantages (e.g. backed artefacts), ceremonial advantage or, for later stages of the expansion, the dingo.”
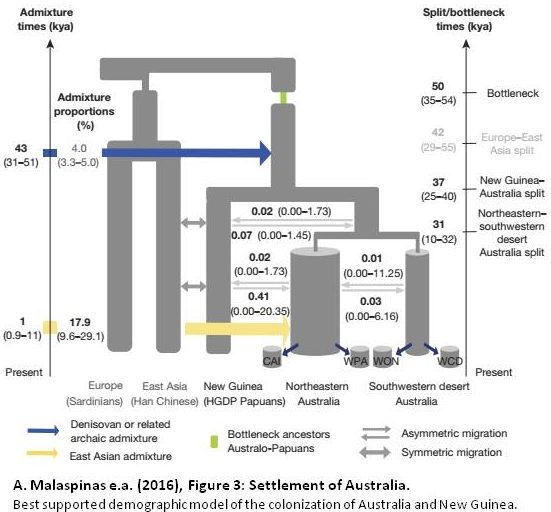 Importantly, this wide-ranging and rapid language replacement seems to have hardly involved genetic change. A. Malaspinas e.a. (2016) “A genomic history of Aboriginal Australia” suggest a split between NE and SW Australian populations already some 30,000 years ago, with very little genetic exchange afterwards. Malaspinas points at climate-induced bottlenecks in SW Australia, and a NE Australian population expansion during the Holocene (last 10,000 years). And, in their Supplementary Notes, Bouckaert e.a. present a number of cases where the archaeological record shows recent resettlement that correlates with their inferred separation time of Pama-Nyungan subgroups. Nevertheless, parts of South and SW Australia have been continuously settled for at least 20,000 years, and apparently relatively recently shifted to Pama-Nyungan without involvement of substantial geneflow. That is a remarkable finding, even more so when considering that Pama-Nyungan speakers were hunter-gatherers.
Importantly, this wide-ranging and rapid language replacement seems to have hardly involved genetic change. A. Malaspinas e.a. (2016) “A genomic history of Aboriginal Australia” suggest a split between NE and SW Australian populations already some 30,000 years ago, with very little genetic exchange afterwards. Malaspinas points at climate-induced bottlenecks in SW Australia, and a NE Australian population expansion during the Holocene (last 10,000 years). And, in their Supplementary Notes, Bouckaert e.a. present a number of cases where the archaeological record shows recent resettlement that correlates with their inferred separation time of Pama-Nyungan subgroups. Nevertheless, parts of South and SW Australia have been continuously settled for at least 20,000 years, and apparently relatively recently shifted to Pama-Nyungan without involvement of substantial geneflow. That is a remarkable finding, even more so when considering that Pama-Nyungan speakers were hunter-gatherers.
Lets move across the Pacific to NW Canada: Coastal Tsimhians, settling around Prince Rupert near the border between British Columbia and Alaska, are among the best studied Native American Groups, thanks not only to analysis of modern DNA, but also of 25 ancient genomes from the region dating to ∼6,260 – 1,036 cal BP, with most of the individuals falling between 3,000 and 1,036 cal BP. John Lindo e.a.(2016) have on the Tsimshians done “A time transect of exomes from a Native American population before and after European contact“, that was extended in a 2018 follow-up paper headed by the same researcher. In a further paper (Lindo e.a. 2016b), the ancient genomes are assessed for their relation to other North American ancients (Anzick-1, Kennewick, etc,) as well as to native North- and South Americans.
My feeling is that the late Pleistocene and early Holocene of the Pacific NW requires further studying. Lindo e.a. 2016b report “that population structure existed in the late Pleistocene of North America“, and “regional shifts in mitochondrial DNA haplogroups“. The latter, more specifically, means disappearance of mtDNA hg D4h3a from the N. American record after ca, 4,000 BC, while it is still present in modern native S. Americans. Such complexity and structure would of course better explain the NW American pre-European linguistic patchwork than the standard “single founding population, local continuity” meme commonly brought forward. However, that is an issue deserving a post on its own.
Wherever Tsimshians ultimately originated from (they are being connected to the somewhat controversial Penutian languages from California and Oregon), they display genetic continuity with local ancient genomes dating back to ca. 3,600 BC, and archeology suggests at least 5,000 years of settlement continuity. Lindo e.a. 2018 report “evidence that the indigenous population was on a steady decline in effective population size for several thousand years before [European] contact“, an issue unfortunately not explored in detail that IMO may have to do with the Athabsacan (Na Dene) southwards expansion. A major demographic shock occured after European contact some 175 years ago, when effective population size fell by some 57%. This aligns well with the historical record of Tsimshian population decreasing from ca. 8,500 in 1835 to around 4,000 by the end of the 19th century a/o due to several smallpox epidemics, the largest of which occured in 1862. [If you think 57% is a huge death toll, compare this fresh paper by said J. Lindo that infers over 90% population collapse for lowland Andean Huilliche-Pehuenche following European contact. The Tahitian population is estimated to have decreased by more than 80% after having been visited by Bougainville, James Cook, and the Bounty.]
Subsequently, Coastal Tsimshian population numbers recovered somewhat in a process of heavy admixture with Europeans, but also surrounding Native Americans, to now in Canada amount to 5,910 (self-declared, 2016 census). European admixture in Coastal Tsimshians, while not present even in the youngest of the ancient genomes, currently stands around 30% (Lindo 2016, 2018), with a strongly sex-biased pattern. A recent study (Hughes e.a. 2017) reports 0% European mtDNA vs, 63% European yDNA (37% R1b, 22% I1/I2b1, 4% G2a) among Coastal Tsimshians.
Admittedly, this isn’t the best example to be put forward – just the best documented case we have from the Americas:
- Male driven-genetic replacement, while substantial at 63%, isn’t overwhelming; and
- At 410 Canadian speakers of Coastal Tsimshian (2,730 speakers of Tsimshian languages overall, which probably entails substantial double counting) according to the 2016 Census, Coastal Tsimshian is seriously endangered of extinction.
Nevertheless, one needs to consider that Tsimshians, and Tsimshian languages, have been under pressure long before European arrival by the Athabascan epansion from the NE, and the Algic-Wakashan-Salish Sprachbund to the south (more on the latter in a separate post). Moreover, the Port Rupert area has seen substantial European colonisation, a/o linked to the Yukon goldrush, turning Coastal Tsimshians into a demographic minority in their traditional settlement area. Last but not least, Tsimshians apparently have always been a relatively small group number-wise. Against this background, the fact that Coastal Tsimshian, however endangered, has so far managed to evade extinction is IMO already remarkable – there are manifold examples of NW Pacific Native American languages with a worse fate.
What may explain the resilience of Coastal Tsimshian, in spite of substantial male-driven genetic replacement, and European colonisation of the traditional settlement area? Clearly, Tsimshian isn’t a lingua franca, and most likely also wasn’t one in pre-European times (that function should rather have fallen on either Athabascan or Wakashan-Salish). However, as common along most of the NW Pacific coast (and also characteristic of the “Pueblo Cultures” of the SW USA), Tsimshian culture is matriarchic / matrilocal.
Further east, around and NE of the Great Lakes, we find the Algonquian-speaking Ojibwe (Chippewa). Ojibwayan languages are considered to be one of the most healthiest Native American language groups in N. America, with some 100,000 speakers acc. to Wikipedia. The 2016 Canadian Census reports nearly 19,000 people still speaking Ojibwayan most often at home, and the US, which I didn’t look up for figures, should add further to that number.
At the same time, the Ojibwe are famous for their high proportion of yDNA R1(b) that, depending on the study author, may range up to 79%. Most of the studies come from the mid-2000s, and we could certainly do well with more recent analyses that consider further hgs of presumably European origin such as I and J. In addition, while contact with 17th century French missionaries, explorers and fur traders is attested, earlier contact with Vikings/ Icelanders, either sailing down the St. Lawrence River, or exploring the Hudson Bay, is also imaginable, and DNA data might help in confirming or rejecting such contact.
In summary: The Ojibwe appear to represent a case of substantial male-driven replacement of Amerindian by European DNA, the detailed mechanism of which still needs to be established, that didn’t trigger language shift. As to “why not?”, matriarchy here doesn’t provide an explanation, as Ojibwe are described as being patriarchic. Howewer, they appear to have a specific “immigration control” mechanism, whereby anybody with non-Ojibwe descent needs to be adopted by a tribe member to be accepted within the tribe. Otherwise, of course, Algic was once spoken from California to Newfoundland, and as such clearly served as lingua franca.
Let me finish the journey around the world (in what seems to come close to 80k letters) in the Levante. Haber e.a. (2017) have presented aDNA from BA Sidon (ca. 1,700 BC), possibly the earliest West Eurasian sample with clear linguistic afiliation, namely to NW-Semitic (pre-Phoenician). Lazarides e.a. (2016) have presented further Levante_BA samples from Jordan, possibly a bit older but with a less clear linguistic assignment. All four male samples presented by Haber e.a. and Lazarides e.a. had yDNA J, signifying a clear break with Natufians and Levantine_N that, to the extent samples could be assigned to a specific yDNA hg beyond basal CT, carried E1b1, H2 and T. That break is still visible in modern populations – acc. to Eupedia, Levante_N yDNA (E, T) just makes up for 22% of Lebanese, and 27% of Palestinian yDNA, that are both dominated by yDNA J.
While having been found in a singleton Anatolian farmer, an EHG from Karelia and IIRC also some EEF, yDNA J is earliest and best attested from EP and Mesolithic contexts around the Caucasus, namely Satsurblia (CHG, J1), Hotu Cave (Iran_HG, J2) and Kotias (CHG, J2). Does that now mean that CHG/ Iran_HG spoke proto-AfroAsiatic (pre-proto Semitic)? Of, course, not!
- Haber e.a. have estimated the Sidon_BA population to be an admixture of 52% Iran_Chl and 48% Levant_N (which suggests, when looking at ancient and modern yDNA, a primarily male-driven process of genetic replacement). Notably, their f4-statistics yield a much stronger admixture signal for CHG than for Iran_Chl, but such scenario wasn’t explored further.
- From linkage equilibrium, they have estimated the admixture date at around 3,000 BC. This fits well with the arrival of the Kura-Araxes culture in the Levante, where it formed the Khirbet-Kerak Culture. We don’t have KA yDNA so far (Areni Cave Armenia_CA may have rather represented Chaff-faced Ware), but connection of KA to yDNA J looks anything but unlikely.
- Arrival around 3,000 BC is clearly too late for having spread Proto-Afro-Asiatic, as it is contemporary with the earliest attestations of Old Egyptian that by that time was already fairly differentiated from Proto-Semitic. It also seems too late for Proto-Semitic, commonly dated to the early 4th mBC.
- Moreover, innovations in Old Akkadian point to a spread from West to East. Several specifics of Old Akkadian, such as the SOV word order or loss of most fricatives are attributed to Sumerian influence. If Proto-Semitic had originated near the Caucasus, and spread from there to the Levante, it should have incorporated said contact phenomena with Sumerian, so they would have become characteristic of Semitic as a whole and not just of Akkadian.
The questions of the Proto-AfroAsiatic homeland, and of uniparental markers possibly linked to the family, deserves a posting of its own. Let me just say here that
- the recently established genetic connection of Iberomaurusians and Natufians opens up intriguing new linguistic possibilities, namely Afroasiatic at least in parts reflecting Paleolithic substrate shared by Berbers, Old Egyptians and Semites, rather than their languages being directly related, and
- certain homeland theories, such as the Horn of Africa, seem untenable anymore in view of the obvious lack of Mota-like, and more generally Sub-Saharan ancestry in Canaans (and already Natufians, see Lazarides e.a. 2016).
I personally tend towards a strong link between yDNA E1b1 and Afro-Asiatic, but that is admittedly speculative.
Anyway – back to the main point: Apparently, there was massive migration from the Caucasus/ Zagros into the Levante around 3,000 BC that substanially shifted the autosomal and even more so the yDNA profile of the region. That shift can plausibly be attributed to the Kura-Araxes expansion. Nevertheless, the immigrants adapted Semitic instead of imposing their language. Why? The CA/EBA Levante doesn’t really present itself as matriarchic (though there is some hypothising on matriarchic structures on Minoan Crete, and Cleopatra signifies certain matriarchic Egyptian traditions). OTOH, KA fits well into the paradigm of a semi-nomadic, pastoralist and ultimately patriarchic society. This leaves the issue of lingua franca; With the powerful, Semitic-speaking Akkadian empire to the NE, and the equally powerful, Afro-Asiatic speaking Egypt to the SW, it is certainly not a bad idea to adapt a local dialect that may be understandable to both of them …
Bottom lines:
- Male-driven genetic replacement without language change appears to have been more frequent than commonly assumed. I have presented here three cases relating to major language families in terms of geographical spread, namely Austronesian, Afro-Asiatic, and Algic. “Uralic” N1c may become case #4.
- Gender-relations of the “giving” and “receiving” groups seem to play an important role. Matriarchic/ matrilocal societies, often found on coasts, appear to be relatively resilient to male-induced language shift. The other side of the spectrum is represented by (semi-) nomadic pastoralists with typically patriarchic structures, who, at least across most of Africa, could quickly impose their yDNA and language on other populations encountered.
- Most important, however, seems to be the issue of lingua franca, i.e. speaking (and possibly adapting) a language that neighbouring (potential) trade partners can understand. Australian Pama-Nyungan languages demonstrate that mutual understandability isn’t just relevant for modern societies, but apparently already played an important role in HG contexts. Unsurprisingly, the same applies to the Near Eastern CA/BA with its long-distance trade relations.
Stll going through this but the native languages of pre-colonial New World can inform us a lot about prehistoric West Eurasia, as has long been recognised (but of course they should be studied in their own right, and not merely as a tool for analogizing 😉 ).
The Amerindian languages beautifully illustrate how rapid language divergence can be – one or two (or whichever the current stance is) migrations into North America, with linguistic bottlenecking in Beringia, yet thousands of years later we have languages which no longer form one or two macro -families. This must mean that as low-density hunter-gatherers expand into new ecological niches, ”specialize” locally and break former contacts, marked language divergence is the rule, with a variegated langoscape (barring the much later arriving, more expansive Na-Dene).
We can draw some broad analogies to Europe and Siberia, after the LGM as new lands were (re-)populated from south but also latitudinal trans-migrations of microblade peoples.
Thousands of years later, as the EEFs migrated through Europe and moved beyond the loess, & engaged in regionally specific adaptation and admixture, whilst at the same time experience ongoing migratory flow from the east and southeast, whatever EEF language that initially existed must have become broken up into several parts by the Late Neolithic.
About Pama-Nyungan, it had long attracted attention because it runs counter to the Bellwood-Renfrew hypothesis: that the most common mechanism of language spread in pre-state times muct have been demography-subsistence based (e.g. farming). However, from what little I’ve read, the expansion of P-N certainly carried a demic component (often moving into empty areas “”downstreamm” migration) as well as a ”subsistence” one (the new backed blades for hunting, presumably).
Of course, we now know that EEFs weren’t the only large migration through Europe, the steppe migration was large – at least through northern and western Europe. Yet this was a subsistence-driven one too, at least as far as the sub-Balticum and central Asia goes, because it represents the expansion of a productive economy in zones still ‘sub-Neolithic’ in economy.
Just sharing my anecdotal experience. Being born in Africa, in spite of being very young at the time , I do remember the following: Males moving in tribal regions always seemed to immediately try to pick-up and learn the local languages, at least to engage and be able to communicate (there are a lot of Portuguese old man that still speak some kimbundo and Umbundo words). One would think it were mostly, if not just, men doing the first contacts with different population over pre-history. So, as in Colonial Africa, immediately started the changes in Y-Dna in local dna, but not at all language. There were from the firs generation in some villages 5%-10% turnover in Y-Dna with absolutely no impact on language. It was the arrival of “Family package” that seems to “impose” language. – Its always with strong cultural traits that you see the shift in local populations to the language of the incoming.
– Any of you guys know more recent examples were it did not happen this way?
Heck even I remember some words: Wakolopo, ovangula umbundu Angola.
I guess we are all basically in agreement that there’s not a single rule in language transmission and that each case needs to be analysed on its own circumstances. Lack of disagreement about this main argument is probably why there’s not a lot of debate going on here.
However, this was a necessary post for setting some basics for upcoming and more complicated debates more centred on the basic scope of this blog.
The most interesting case within this scope discussed above is the one about the NW Iran/Caucasus migrations to the Levant. While there probably was a good amount of continuous flow (as seen already in the Chalcolithic samples), it’s quite well documented archaeologically and now supported by aDNA the Bronze Age incursion of the Kura-Araxes people into the region. And indeed, it seems unlikely that they brought Semitic languages with them. So the role of a well established local language, specially if widespread already and that could serve as Lingua Franca is one of the factors to take into account when looking at possible language shift.
Also what Olympus Mons says about the cultural and “family” package is a very important factor, more so than male biased migration alone. Also the “organized state” package (as in the language shift in Iberia with the Roman presence).
I hope that all these things will come up when we get (when Frank and I sort out how to deal with it) to Western Europe and the possible language affiliation of Neolithic Farmers, Bell Beakers, the Celtic expansion, etc…
But now I guess it’s time to look at those Siberian samples. Hope to be able to write something up in the next couple of days.
OM (welcome here, btw):
Equally anecdotic – my grasp of South Slavic significantly benefitted from a relatively short-lived affair with a Bulgarian girl back in my youth (mid- 1990s).
As to other examples: I tried to be quite comprehensive (maybe too much, considering how long the post became) as concerns cases with clear genetic evidence. East Africa, especially Madagascar, is an area certainly deserving more in-depth research. Parts of India today speaking IE languages with strong Dravidian linguistic substrate also come to mind, but the absence of ancient aDNA from the subcontinent, and the uncertainty of when and how both IE and Dravidian reached the subcontinent at the moment prevents any substantiated analysis.
The main idea of the post was of course related to Europe, and more specifically towards sending some warning against automatically equalling the male-driven advance of “Steppe” ancestry with the spread of IE. Especially Basque, a non-IE language spoken in an area that apparently experienced substantial male-driven replacement during the Bell Beaker period, is conspicious in this respect, but I will leave it to Alberto to address that question in more detail whenever he sees it fit (naturally, a lot of related literature is in Spanish and as such better understandably to Alberto than to myself as a German).
Alberto: You mentionned the Lazarides 2016 Levante_CA samples as already demonstrating CHG migrations into the Levante. Note that these samples equally post-date the Kura-Araxes (Khirbet-Kerak ware) expansion.
There was an earlier expansion out of (or at least linked to) the Caucasus, namely that of chaff-faced ware (CFW), a/o connected to Leila Tepe, Sioni and Maykop, but archeology suggests that CWF only made it to the Northern Levante, i.e. the Antakhyia Region, during the 1st half of the 4th mBC. That of course neither precludes that CWF already carried CHG/ Iran_N ancestry to the N. Levante, nor that this ancestry subsequently trickled down further south – to the opposite, both is quite likely. Nevertheless, as concerns Lebanese / Palestinians, IMO the main agent seems to have been KA. Ultimately, we need more ancient DNA from the region for certainty.
There is one general point that I intended to make but didn’t spell out clearly in my post: Male-driven genetic replacement isn’t generally meaning “kill the males, rape the women”. We know that this has happened – not so long ago in the Bosnian Civil War, there is the mythological Roman “Rape of the Sabine women”, and we have evidence of violently destroyed LBK settlements with a suspicious lack of female corpses of reproductive age among the victims. So, such a scenario is certainly to be envisaged when it comes to male-driven genetic replacement.
But there are at least two other possible scenarios:
1. A cultural pre-disposition to search for external mating partners outside the own tribal (ethnic) group. Such cultural pre-disposition is evolutionary favorable, as it reduces the risk of incest-related offspring damage, especially if the tribal group in question is numerically small. Moreover, a minimal headcount of around 1,000 is required for a population to survive. As has been demonstrated by stochastic modeling, at lower headcounts there will develop a gender imbalance that over the course of a few generations will result in extinction. As such, if you don’t “trade off” your excess girls or boys, respectively, to a neighboring group, you will be quickly done. [Rob, btw: This is a major reason why I don’t think that HG languages could diversify substantially. Being small in headcount, HG groups needed genetic exchange with neighboring groups, and that requires the ability for mutual communication.]
Such cultural pre-disposition is well documented for Polynesia, c.f. the “Mutinity on the Bounty”, and has inspired lots of 19th/ early 20th century phantasies about a South Pacific “paradise of free love”. Given Polynesia’s insular nature, of course, we are not dealing with some utopian philosophy here, but with evolutionally rational behavior. Such behavior most likely also explains how Papuan male ancestry came to replace the original Austronesian one. Matriarchy looks like a plausible corrective to ensure that, for all the external admixture, cultural coherence on the island/ within the tribe can be maintained.
2. Massive disease- related demographic shock, as experienced, e.g., by Tsimshians. That’s a complex scenario. It first of all means contact with another, previously unencountered, expanding population that transmits the disease. Secondly, the demographic shock allows for various scenarios of genetic shift, including colonization of depopulated areas by that expanding population – a scenario that might be envisaged for EEF, which certainly had acquired better resistance against diseases linked to animal husbandry such as smallpox, measles and tuberculosis, than WHG, and also for the “Steppe” expansion (here, the issue is the Plague). However, colonization isn’t male-driven genetic replacement, so how does the gender imbalance fit into here?
While the issue is far from being fully understood, there is quite some evidence that mtDNA conveys different degrees of resistance to specific diseases. The same may also apply to yDNA. E.g., available records indicate that the medieval Black Death caused the highest death toll in Tuscany (yDNA E, J), and in E. England, N. Germany, and Scandinavia (yDNA I). Conversely, areas such as Cornwall / Wales, the Basque Country, and Central Europe between Nuremberg and Cracow (all heavy in R1a/b) seem to have been much less or hardly affected at all. So, the demographic shock may have been gender-specific, killing more males with a specific yDNA than females overall.
Even if an epidemic decimates males and females alike, a reasonable reaction is to seek for external mating partners, as potential mating partners from the same tribe have become rare. This applies even more to populations with a cultural pre-disposition to search for external mating partners as described above.
Returning to one of the cases discussed in my post, namely the Ojibwe: They certainly have a quite long history of interaction with Europeans, going back to French and Hudson Bay Company fur traders, possibly even to Green-/ Icelanders. However, I haven’t come across anything suggesting a “kill the males, rape the women” scenario. Certainly not from the French-Canadian side – the Ojibwe were part of the Wabanaki Confederacy that fought alongside the French in the 1754-63 French and Indian War. As the Wabanaki Confederacy afterwards sided with American revolutionaries against Britain, and therefore was always considered “friendly” by US governments, there is also little reason to suppose such a scenario for the US colonization of the Mid-West.